Krautschau: Was wächst zwischen den Pflasterfugen Deiner Stadt?
Bundesweite Aktionswoche vom 18. bis 26. Mai 2024
Die oft übersehenen kleinen Pflanzen in Pflasterfugen und Mauerritzen sind die geheimen Stars der städtischen Flora. mehr
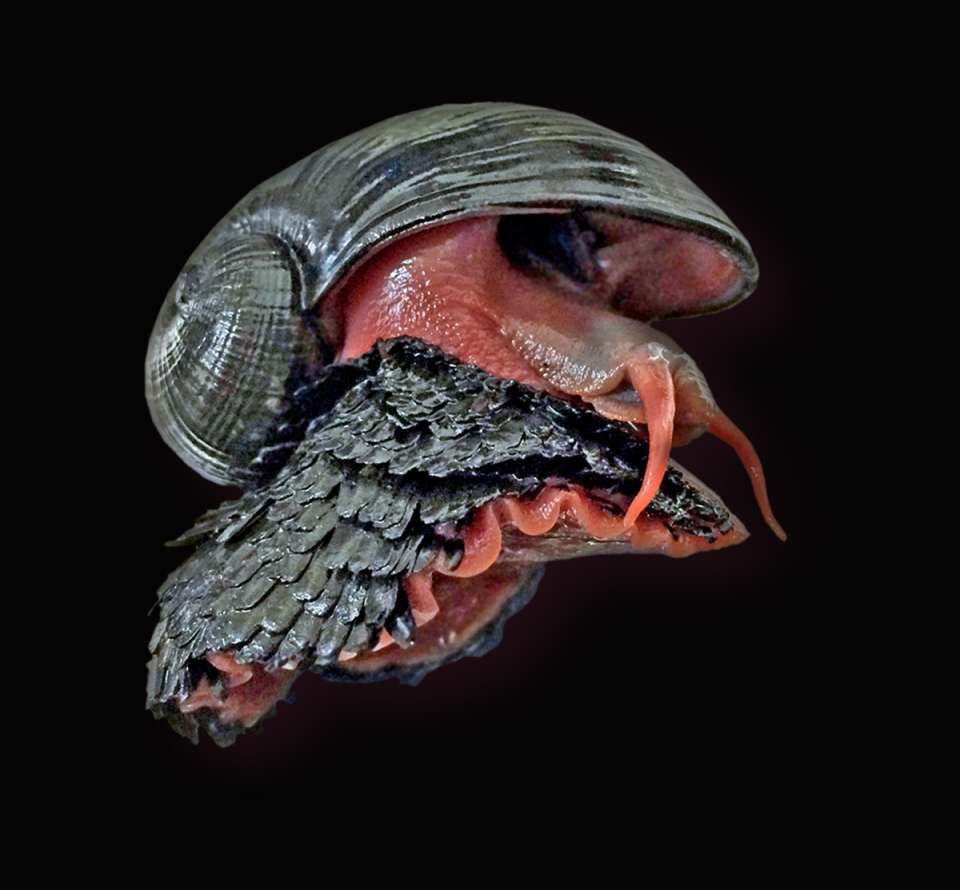
Maskierte Räuber: Waschbären sind eine Gefahr für heimische Amphibien und Reptilien
Untersuchung in Naturschutzgebieten in Hessen, Sachsen-Anhalt und Brandenburg durchgeführt
Der Frankfurter Parasitologe und Infektionsbiologe Prof. Dr. Sven Klimpel zeigt gemeinsam mit weiteren Forschenden im Rahmen des Verbundprojektes ZOWIAC in einer aktuellen wissenschaftlichen Studie, dass Waschbären in Deutschland Amphibien und Reptilien als Nahrungsquelle nutzen. mehr
Senckenberg-Plakatkampagne in Frankfurt macht neugierig auf Meeresbewohner
Vier Meeresbewohner, vier Sprüche, ein Ziel: Seit heute zeigt das Senckenberg Forschungsinstitut und Naturmuseum Frankfurt im Rahmen einer Plakatkampagne in der Mainmetropole wirbellose Kleinstlebewesen aus dem Meer. mehr
Senckenberg To Go – Auf dem Weg in die Arbeit und zur Schule oder ganz bequem von Zuhause. Senckenberg bietet Ihnen verschiedene unterhaltsame und informative Online-Formate mit denen sie ihr Wissen über unseren Planeten mühelos erweitern können.
Bei einem außergewöhnlichen Waldspaziergang durch die Sonderausstellung im Senckenberg Naturmuseum eröffnen sich Einblicke in das Ökosystem Wald. Waldbilder und -vorstellungen aus der Romantik treffen auf naturwissenschaftliche Forschung zur Evolution von Wäldern und deren Zukunft. Welche Rolle spielen Wälder für uns und wie ist unser Verhältnis zu Wäldern geprägt? Der Weg führt uns unter anderem zu einer indigenen Universität des Waldwissens im Amazonasgebiet, einem Protestcamp zum Waldsterben bis hin zu einem Kameraflug von den Wurzeln in die Wipfel eines virtuellen Urwalds.
Kosten: 5,- Euro pro Person, zzgl. Eintritt
Tickets erhalten Sie online oder nach Verfügbarkeit auch an der Museumskasse
Bitte beachten Sie: das Ticket beinhaltet keinen Eintritt ins Museum. Dieser muss extra gebucht werden. Teilnahme nur mit Eintrittsticket & Führungsticket möglich
Dauer: 1 Stunde Treffpunkt: Senckenberg Naturmuseum, am Anfang der Sonderausstellung, im Vogelsaal, 1.OG
Führung für Erwachsene zur Sonderausstellung Wälder. Von der Romantik in die Zukunft
Wälder. Von der Romantik in die Zukunft
Onlineticket
Erwachsene
Jugendliche
Bei einem außergewöhnlichen Waldspaziergang durch die Sonderausstellung im Senckenberg Naturmuseum eröffnen sich Einblicke in das Ökosystem Wald. Waldbilder und -vorstellungen aus der Romantik treffen auf naturwissenschaftliche Forschung zur Evolution von Wäldern und deren Zukunft. Welche Rolle spielen Wälder für uns und wie ist unser Verhältnis zu Wäldern geprägt? Der Weg führt uns unter anderem zu einer indigenen Universität des Waldwissens im Amazonasgebiet, einem Protestcamp zum Waldsterben bis hin zu einem Kameraflug von den Wurzeln in die Wipfel eines virtuellen Urwalds.
Kosten: 5,- Euro pro Person, zzgl. Eintritt
Tickets erhalten Sie online oder nach Verfügbarkeit auch an der Museumskasse
Bitte beachten Sie: das Ticket beinhaltet keinen Eintritt ins Museum. Dieser muss extra gebucht werden. Teilnahme nur mit Eintrittsticket & Führungsticket möglich
Dauer: 1 Stunde Treffpunkt: Senckenberg Naturmuseum, am Anfang der Sonderausstellung, im Vogelsaal, 1.OG
Diese Exkursionen bieten eine einzigartige Gelegenheit, die oft übersehene Schönheit unserer urbanen Umgebung zu entdecken und schätzen zu lernen. Interessierte sind herzlich eingeladen, an diesen Exkursionen teilzunehmen und die unscheinbaren Schätze unserer urbanen Umgebung zu entdecken.
Die Teilnahme ist kostenlos und dauert ca. 2 Stunden.
Diese Exkursionen bieten eine einzigartige Gelegenheit, die oft übersehene Schönheit unserer urbanen Umgebung zu entdecken und schätzen zu lernen. Interessierte sind herzlich eingeladen, an diesen Exkursionen teilzunehmen und die unscheinbaren Schätze unserer urbanen Umgebung zu entdecken.
Die Teilnahme ist kostenlos und dauert ca. 2 Stunden.
Zur Sonderausstellung „Wälder. Von der Romantik in die Zukunft“ kooperiert die Dresden Frankfurt Dance Company mit dem Senckenberg Naturmuseum und bietet im Rahmen der Ausstellung zu ausgewählten Terminen eine Performance an, die Besuchende dazu einlädt, sich gemeinsam auf eine Reise durch den Wald zu machen und darin zu verlaufen. Während sie eine Tänzerin oder einen Tänzer der Dresden Frankfurt Dance Company auf deren Weg durch die Räume des Senckenberg Naturmuseums begleiten, machen alle ihre je eigene Reise. Dabei hören die Besucher*innen eine Tonspur, die das Geschehen emotional rahmt. Die Choreografie verschiebt die Aufmerksamkeit von den Ausstellungsstücken zu den tanzenden Körpern, die sich im Wald der Ausstellung verlieren. Indem man die Umgebung im Augenwinkel wahrnimmt, verwandelt sich der Raum zu einem Ort, der mehr zu sagen hat, als die Informationen, die in ihm aufbereitet sind. Er wird zum Schauplatz einer Reise, deren Rätsel man nicht ganz entschlüsseln kann. Wo bin ich jetzt? Wo bin ich hingeraten? Was ist das für ein Ort?
ab 18:30 Uhr, jeweils drei Zeit-Slots im 20-Minuten-Takt
Im Kino des Senckenberg Naturmuseums sind über die Laufzeit der Ausstellung Filmaufnahmen der Tanzproben zu „If you don’t get lost in the woods, you haven’t been to the woods“ jeweils zu den Museumsöffnungszeiten und immer zur vollen Stunde zu sehen.
Video mit Ton und Untertiteln, 22:22 Min.
Choreographie: Ioannis Mandafounis, Dresden Frankfurt Dance Company
Tänzer:innen: Nastia Ivanova und Thomas Bradley, Dresden Frankfurt Dance Company
Film, Ton und Schnitt: Nicklas Bauske und Adrian Giacomelli, Senckenberg Gesellschaft für Naturforschung
If you don’t get lost in the woods, you haven’t been to the woods
Tanz-Performance mit Tänzer*innen der Dresden Frankfurt Dance Company
Wälder. Von der Romantik in die Zukunft
Onlineticket
Erwachsene
Jugendliche
Foyer
eine Choreographie von Ioannis Mandafounis
Zur Sonderausstellung „Wälder. Von der Romantik in die Zukunft“ kooperiert die Dresden Frankfurt Dance Company mit dem Senckenberg Naturmuseum und bietet im Rahmen der Ausstellung zu ausgewählten Terminen eine Performance an, die Besuchende dazu einlädt, sich gemeinsam auf eine Reise durch den Wald zu machen und darin zu verlaufen. Während sie eine Tänzerin oder einen Tänzer der Dresden Frankfurt Dance Company auf deren Weg durch die Räume des Senckenberg Naturmuseums begleiten, machen alle ihre je eigene Reise. Dabei hören die Besucher*innen eine Tonspur, die das Geschehen emotional rahmt. Die Choreografie verschiebt die Aufmerksamkeit von den Ausstellungsstücken zu den tanzenden Körpern, die sich im Wald der Ausstellung verlieren. Indem man die Umgebung im Augenwinkel wahrnimmt, verwandelt sich der Raum zu einem Ort, der mehr zu sagen hat, als die Informationen, die in ihm aufbereitet sind. Er wird zum Schauplatz einer Reise, deren Rätsel man nicht ganz entschlüsseln kann. Wo bin ich jetzt? Wo bin ich hingeraten? Was ist das für ein Ort?
ab 18:30 Uhr, jeweils drei Zeit-Slots im 20-Minuten-Takt
Im Kino des Senckenberg Naturmuseums sind über die Laufzeit der Ausstellung Filmaufnahmen der Tanzproben zu „If you don’t get lost in the woods, you haven’t been to the woods“ jeweils zu den Museumsöffnungszeiten und immer zur vollen Stunde zu sehen.
Video mit Ton und Untertiteln, 22:22 Min.
Choreographie: Ioannis Mandafounis, Dresden Frankfurt Dance Company
Tänzer:innen: Nastia Ivanova und Thomas Bradley, Dresden Frankfurt Dance Company
Film, Ton und Schnitt: Nicklas Bauske und Adrian Giacomelli, Senckenberg Gesellschaft für Naturforschung
Wissenschaftler*innen aus ganz Deutschland beantworten für Kinder zwischen 8 und 12 Jahren spannende Fragen aus ihrer Forschung. Wie an einer richtigen Hochschule bekommen die Kinder einen Studierendenausweis und am Ende jedes Semesters ein Diplom. Die Teilnahme an der Kinderakademie ist kostenlos und geschieht auf eigene Gefahr. Eltern sind zu den Vorlesungen nicht zugelassen. Wir bitten um eine Anmeldung für das Semester.
Die Görlitzer Kinderakademie ist eine gemeinsame Veranstaltung des SENCKENBERG Museums für Naturkunde Görlitz und der Hochschule Zittau/Görlitz
Förderer der Kinderakademie: Förderkreis des Naturkundemuseums Görlitz e. V. und Förderverein der Hochschule Zittau/Görlitz e. V.
Vorlesung nur für Kinder | Gastbeitrag der polnischen Hochschulen, Partner im Akademischen Koordinierungszentrum Euroregion Neiße
Anmeldung erforderlich / Registration required
Kinder
38. SEMESTER
Wissenschaftler*innen aus ganz Deutschland beantworten für Kinder zwischen 8 und 12 Jahren spannende Fragen aus ihrer Forschung. Wie an einer richtigen Hochschule bekommen die Kinder einen Studierendenausweis und am Ende jedes Semesters ein Diplom. Die Teilnahme an der Kinderakademie ist kostenlos und geschieht auf eigene Gefahr. Eltern sind zu den Vorlesungen nicht zugelassen. Wir bitten um eine Anmeldung für das Semester.
Die Görlitzer Kinderakademie ist eine gemeinsame Veranstaltung des SENCKENBERG Museums für Naturkunde Görlitz und der Hochschule Zittau/Görlitz
Förderer der Kinderakademie: Förderkreis des Naturkundemuseums Görlitz e. V. und Förderverein der Hochschule Zittau/Görlitz e. V.
Offene Themenstation: Insekten – Vielfalt auf sechs Beinen
Insekten sind eklige und nervige Krabbler? Von wegen! Insekten sind nützliche und
faszinierend schöne Tiere! Unter dem Mikroskop offenbaren sie eine unglaubliche
Farben-und Formenvielfalt, die es zu entdecken gilt. Dazu stehen unsere
Expert*innen Rede und Antwort und verraten, wie man Insekten erkennen,
unterscheiden und im eigenen Garten unterstützen kann.
Keine Anmeldung erforderlich Ab 8 Jahren, Kinder nur in Begleitung eines Erwachsenen Angebot nicht für größere Gruppen geeignet Die Teilnahme ist im Eintrittspreis inbegriffen Dauer: In der Zeit von 15:00 – 17:00 Uhr Ort: Aha?! Forschungswerkstatt
Offene Themenstation: Insekten – Vielfalt auf sechs Beinen
ohne Anmeldung / no Registration
Erwachsene
Jugendliche
Kinder
Familien
Offene Themenstation: Insekten – Vielfalt auf sechs Beinen
Insekten sind eklige und nervige Krabbler? Von wegen! Insekten sind nützliche und
faszinierend schöne Tiere! Unter dem Mikroskop offenbaren sie eine unglaubliche
Farben-und Formenvielfalt, die es zu entdecken gilt. Dazu stehen unsere
Expert*innen Rede und Antwort und verraten, wie man Insekten erkennen,
unterscheiden und im eigenen Garten unterstützen kann.
Keine Anmeldung erforderlich Ab 8 Jahren, Kinder nur in Begleitung eines Erwachsenen Angebot nicht für größere Gruppen geeignet Die Teilnahme ist im Eintrittspreis inbegriffen Dauer: In der Zeit von 15:00 – 17:00 Uhr Ort: Aha?! Forschungswerkstatt
In Kooperation mit dem Botanischen Garten und Palmengarten bieten wir im Rahmen der Sonderausstellung "Wälder. Von der Romantik in die Zukunft" einmal im Monat sonntags Einblicke in die Wälder der Welt. Die Führungen finden jeweils im Botanischen Garten oder Palmengarten statt.
Im Reich der dornig-stacheligen Baumgestalten: Wälder der Trockenen Tropen | mit Dr. Ester Becker
In vielen tropischen Regionen regnet es selten. Waldpflanzen aus Trockengebieten zeigen ganz spezielle Anpassungen, darunter auch die Ausprägung stechender Dornen und spitzer Stacheln. Besonders eindrücklich zeigen das auch die Pflanzen der Dornwälder Madagaskars, denen im Tropicarium fast ein ganzes Schauhaus gewidmet ist.
Keine Anmeldung erforderlich Es fällt Eintritt in den Palmengarten an, die Teilnahme an den Führungen ist im Eintritt inbegriffen Treffpunkt: Im Garten, vor dem Eingangsschauhaus, Palmengarten, Eingang Siesmayerstraße 63, 60323 Frankfurt
Im Reich der dornig-stacheligen Baumgestalten: Wälder der Trockenen Tropen
Führung im Palmengarten
Wälder. Von der Romantik in die Zukunft
ohne Anmeldung / no Registration
Erwachsene
Familien
In Kooperation mit dem Botanischen Garten und Palmengarten bieten wir im Rahmen der Sonderausstellung „Wälder. Von der Romantik in die Zukunft“ einmal im Monat sonntags Einblicke in die Wälder der Welt. Die Führungen finden jeweils im Botanischen Garten oder Palmengarten statt.
Im Reich der dornig-stacheligen Baumgestalten: Wälder der Trockenen Tropen | mit Dr. Ester Becker
In vielen tropischen Regionen regnet es selten. Waldpflanzen aus Trockengebieten zeigen ganz spezielle Anpassungen, darunter auch die Ausprägung stechender Dornen und spitzer Stacheln. Besonders eindrücklich zeigen das auch die Pflanzen der Dornwälder Madagaskars, denen im Tropicarium fast ein ganzes Schauhaus gewidmet ist.
Keine Anmeldung erforderlich Es fällt Eintritt in den Palmengarten an, die Teilnahme an den Führungen ist im Eintritt inbegriffen Treffpunkt: Im Garten, vor dem Eingangsschauhaus, Palmengarten, Eingang Siesmayerstraße 63, 60323 Frankfurt
Bäume und Grünflächen sind Schlüsselelemente für die Anpassung der Städte an den Klimawandel. „Klassische“ Stadtgrün-Elemente wie Parks und Straßenbäume leiden aber selbst stark unter den häufigen Dürresommern. Die sogenannten „Tiny Forests“ hingegen sind eine noch recht neue Form der Stadtbegrünung und gelten als klimaresilienter. Die nach der Methode des japanischen Botanikers Akira Miyawaki gepflanzten naturnahen und standortangepassten Wäldchen zeichnen sich durch ihren geringen Flächenbedarf, schnelles Wachstum und ihren Artenreichtum aus. Zudem werden sie gemeinsam mit der Nachbarschaft geplant und gepflanzt, sie sind ein im besten Sinne des Wortes verbindendes Element in der zukunftsfähigen Stadt.
Der Themenabend beschäftigt sich mit dem Wald in der Stadt, der neuen Miyawaki-Methode und stellt das „MainWäldchen“ und seinen Planungsprozess vor. Das MainWäldchen ist Frankfurts erster Tiny Forest auf öffentlichem Grund und wurde im Dezember 2023 in Eckenheim gepflanzt.
Themenabend "Stadtverwaldung" - Kleine Wäldchen als Baustein für die Stadt im Klimawandel
Wälder. Von der Romantik in die Zukunft
ohne Anmeldung / no Registration
Erwachsene
Mitglieder
Wissenschaftliches Fachpublikum
Schulklassen
Familien
Presse
Grüner Hörsaal
Bäume und Grünflächen sind Schlüsselelemente für die Anpassung der Städte an den Klimawandel. „Klassische“ Stadtgrün-Elemente wie Parks und Straßenbäume leiden aber selbst stark unter den häufigen Dürresommern. Die sogenannten „Tiny Forests“ hingegen sind eine noch recht neue Form der Stadtbegrünung und gelten als klimaresilienter. Die nach der Methode des japanischen Botanikers Akira Miyawaki gepflanzten naturnahen und standortangepassten Wäldchen zeichnen sich durch ihren geringen Flächenbedarf, schnelles Wachstum und ihren Artenreichtum aus. Zudem werden sie gemeinsam mit der Nachbarschaft geplant und gepflanzt, sie sind ein im besten Sinne des Wortes verbindendes Element in der zukunftsfähigen Stadt.
Der Themenabend beschäftigt sich mit dem Wald in der Stadt, der neuen Miyawaki-Methode und stellt das „MainWäldchen“ und seinen Planungsprozess vor. Das MainWäldchen ist Frankfurts erster Tiny Forest auf öffentlichem Grund und wurde im Dezember 2023 in Eckenheim gepflanzt.
Kuratorinnenführung: „Wälder. Von der Romantik in die Zukunft“ | mit Museumsdirektorin
Dr. Brigitte Franzen Heute lädt Frau Dr. Brigitte Franzen zu einem außergewöhnlichen Waldspaziergang durch die Sonderausstellung „Wälder. Von der Romantik in die Zukunft“ ein und erläutert dabei die transdisziplinären Verbindungen. Vor dem Hintergrund von Klima- und Biodiversitätskrisen bringt die Ausstellung am Beispiel des hochromantischen Themas „Wald“ frühe Ansätze zur Entwicklung anderer Naturverhältnisse in Dialog mit aktuellen Fragestellungen und Forschungen. Der Weg führt unter anderem zu einer indigenen Universität des Waldwissens im Amazonasgebiet, einem Protestcamp zum Waldsterben bis hin zu einem Kameraflug von den Wurzeln in die Wipfel eines virtuellen Urwalds.
Keine Anmeldung erforderlich
Die Teilnahme ist im Eintrittspreis inbegriffen
Dauer: ca. 50 Minuten
Treffpunkt: Afrikanischer Elefant im Saal der Wale und Elefanten
Kuratorinnenführung: „Wälder. Von der Romantik in die Zukunft“ | mit Museumsdirektorin Dr. Brigitte Franzen
Wissenschaftliche Themenführung am Mittwochabend für Erwachsene
ohne Anmeldung / no Registration
Erwachsene
Jugendliche
Kuratorinnenführung: „Wälder. Von der Romantik in die Zukunft“ | mit Museumsdirektorin
Dr. Brigitte Franzen Heute lädt Frau Dr. Brigitte Franzen zu einem außergewöhnlichen Waldspaziergang durch die Sonderausstellung „Wälder. Von der Romantik in die Zukunft“ ein und erläutert dabei die transdisziplinären Verbindungen. Vor dem Hintergrund von Klima- und Biodiversitätskrisen bringt die Ausstellung am Beispiel des hochromantischen Themas „Wald“ frühe Ansätze zur Entwicklung anderer Naturverhältnisse in Dialog mit aktuellen Fragestellungen und Forschungen. Der Weg führt unter anderem zu einer indigenen Universität des Waldwissens im Amazonasgebiet, einem Protestcamp zum Waldsterben bis hin zu einem Kameraflug von den Wurzeln in die Wipfel eines virtuellen Urwalds.
Keine Anmeldung erforderlich
Die Teilnahme ist im Eintrittspreis inbegriffen
Dauer: ca. 50 Minuten
Treffpunkt: Afrikanischer Elefant im Saal der Wale und Elefanten
Zur Sonderausstellung „Wälder. Von der Romantik in die Zukunft“ kooperiert die Dresden Frankfurt Dance Company mit dem Senckenberg Naturmuseum und bietet im Rahmen der Ausstellung zu ausgewählten Terminen eine Performance an, die Besuchende dazu einlädt, sich gemeinsam auf eine Reise durch den Wald zu machen und darin zu verlaufen. Während sie eine Tänzerin oder einen Tänzer der Dresden Frankfurt Dance Company auf deren Weg durch die Räume des Senckenberg Naturmuseums begleiten, machen alle ihre je eigene Reise. Dabei hören die Besucher*innen eine Tonspur, die das Geschehen emotional rahmt. Die Choreografie verschiebt die Aufmerksamkeit von den Ausstellungsstücken zu den tanzenden Körpern, die sich im Wald der Ausstellung verlieren. Indem man die Umgebung im Augenwinkel wahrnimmt, verwandelt sich der Raum zu einem Ort, der mehr zu sagen hat, als die Informationen, die in ihm aufbereitet sind. Er wird zum Schauplatz einer Reise, deren Rätsel man nicht ganz entschlüsseln kann. Wo bin ich jetzt? Wo bin ich hingeraten? Was ist das für ein Ort?
ab 18:30 Uhr, jeweils drei Zeit-Slots im 20-Minuten-Takt
Im Kino des Senckenberg Naturmuseums sind über die Laufzeit der Ausstellung Filmaufnahmen der Tanzproben zu „If you don’t get lost in the woods, you haven’t been to the woods“ jeweils zu den Museumsöffnungszeiten und immer zur vollen Stunde zu sehen.
Video mit Ton und Untertiteln, 22:22 Min.
Choreographie: Ioannis Mandafounis, Dresden Frankfurt Dance Company
Tänzer:innen: Nastia Ivanova und Thomas Bradley, Dresden Frankfurt Dance Company
Film, Ton und Schnitt: Nicklas Bauske und Adrian Giacomelli, Senckenberg Gesellschaft für Naturforschung
If you don’t get lost in the woods, you haven’t been to the woods
Tanz-Performance mit Tänzer*innen der Dresden Frankfurt Dance Company
Wälder. Von der Romantik in die Zukunft
Onlineticket
Erwachsene
Jugendliche
Foyer
eine Choreographie von Ioannis Mandafounis
Zur Sonderausstellung „Wälder. Von der Romantik in die Zukunft“ kooperiert die Dresden Frankfurt Dance Company mit dem Senckenberg Naturmuseum und bietet im Rahmen der Ausstellung zu ausgewählten Terminen eine Performance an, die Besuchende dazu einlädt, sich gemeinsam auf eine Reise durch den Wald zu machen und darin zu verlaufen. Während sie eine Tänzerin oder einen Tänzer der Dresden Frankfurt Dance Company auf deren Weg durch die Räume des Senckenberg Naturmuseums begleiten, machen alle ihre je eigene Reise. Dabei hören die Besucher*innen eine Tonspur, die das Geschehen emotional rahmt. Die Choreografie verschiebt die Aufmerksamkeit von den Ausstellungsstücken zu den tanzenden Körpern, die sich im Wald der Ausstellung verlieren. Indem man die Umgebung im Augenwinkel wahrnimmt, verwandelt sich der Raum zu einem Ort, der mehr zu sagen hat, als die Informationen, die in ihm aufbereitet sind. Er wird zum Schauplatz einer Reise, deren Rätsel man nicht ganz entschlüsseln kann. Wo bin ich jetzt? Wo bin ich hingeraten? Was ist das für ein Ort?
ab 18:30 Uhr, jeweils drei Zeit-Slots im 20-Minuten-Takt
Im Kino des Senckenberg Naturmuseums sind über die Laufzeit der Ausstellung Filmaufnahmen der Tanzproben zu „If you don’t get lost in the woods, you haven’t been to the woods“ jeweils zu den Museumsöffnungszeiten und immer zur vollen Stunde zu sehen.
Video mit Ton und Untertiteln, 22:22 Min.
Choreographie: Ioannis Mandafounis, Dresden Frankfurt Dance Company
Tänzer:innen: Nastia Ivanova und Thomas Bradley, Dresden Frankfurt Dance Company
Film, Ton und Schnitt: Nicklas Bauske und Adrian Giacomelli, Senckenberg Gesellschaft für Naturforschung
Vom Erforschen mit allen Sinnen - Welcome on board Vaka Okeanos
Wir nehmen euch mit auf den Polynesischen Katamaran der Okeanos Stiftung im
Mittelmeer. Jeden Sommer gehen dort Meer-begeisterte Jugendliche an Bord und
leben auf und mit dem Meer. Für Viele ist es das Abenteuer ihres Lebens, wenn sie
beginnen, diesen Lebensraum mit allen Sinnen und auch mit unterschiedlichen
Forschungsmethoden zu erkunden. Mit Planktonnetzen, Hydrophon oder Mikroskop.
Was man im Mittelmeer damit alles entdecken kann, erfahrt ihr heute bei
Wissenschaft live. Wir begleiten die Jugendlichen auf ihrer Reise und erkunden das
Mittelmeer selbst mit verschiedenen Sinnen.
Die Ozean-Botschafter*innen der Jugendinitiative WAVES OF ACTION erzählen
euch in der Aha?! Forscherwerkstatt von ihren Erlebnissen und Begegnungen im
Mittelmeer und warum und wie sie sich für gesunde Meere engagieren!
Keine Anmeldung erforderlich Die Teilnahme ist im Eintrittspreis inbegriffen Für Individualbesuchende (Einzelpersonen, Kleingruppen/Familien), nicht für Großgruppen geeignet Ab 8 Jahren empfohlen, Kinder nehmen in Begleitung von Erwachsenen teil Dauer: in der Zeit von 14:00 bis 16:00 Uhr Ort: Aha?! Forschungswerkstatt
Vom Erforschen mit allen Sinnen - Welcome on board Vaka Okeanos
Wissenschaft Live in der Aha?! Forschungswerkstatt
ohne Anmeldung / no Registration
Erwachsene
Jugendliche
Kinder
Familien
Vom Erforschen mit allen Sinnen – Welcome on board Vaka Okeanos
Wir nehmen euch mit auf den Polynesischen Katamaran der Okeanos Stiftung im
Mittelmeer. Jeden Sommer gehen dort Meer-begeisterte Jugendliche an Bord und
leben auf und mit dem Meer. Für Viele ist es das Abenteuer ihres Lebens, wenn sie
beginnen, diesen Lebensraum mit allen Sinnen und auch mit unterschiedlichen
Forschungsmethoden zu erkunden. Mit Planktonnetzen, Hydrophon oder Mikroskop.
Was man im Mittelmeer damit alles entdecken kann, erfahrt ihr heute bei
Wissenschaft live. Wir begleiten die Jugendlichen auf ihrer Reise und erkunden das
Mittelmeer selbst mit verschiedenen Sinnen.
Die Ozean-Botschafter*innen der Jugendinitiative WAVES OF ACTION erzählen
euch in der Aha?! Forscherwerkstatt von ihren Erlebnissen und Begegnungen im
Mittelmeer und warum und wie sie sich für gesunde Meere engagieren!
Keine Anmeldung erforderlich Die Teilnahme ist im Eintrittspreis inbegriffen Für Individualbesuchende (Einzelpersonen, Kleingruppen/Familien), nicht für Großgruppen geeignet Ab 8 Jahren empfohlen, Kinder nehmen in Begleitung von Erwachsenen teil Dauer: in der Zeit von 14:00 bis 16:00 Uhr Ort: Aha?! Forschungswerkstatt
Mit Ihrer Spende ermöglichen Sie der Senckenberg Gesellschaft Naturforschung zu betreiben und unsere Forschungsergebnisse durch Veröffentlichungen, Ausstellungen, Vermittlungsprojekte und viele weitere Initiativen der Allgemeinheit zugänglich zu machen.